Carbon-based life, or organic life as we know it, revolves around carbon’s unique chemical properties. Carbon atoms have four valence electrons, enabling them to form stable covalent bonds with up to four other atoms, including themselves. This tetravalency allows carbon to create long chains (e.g., hydrocarbons), branched structures, and rings, forming the backbone of biomolecules like proteins, lipids, carbohydrates, and nucleic acids. In aqueous environments—essential for life—carbon compounds exhibit versatility: they can be hydrophobic (repelling water, like fats) or hydrophilic (attracting water, like sugars), facilitating cellular compartments and reactions. Earth’s biochemistry predominantly uses carbon because it’s abundant, lightweight, and forms strong yet flexible bonds, unlike silicon (which forms brittle chains) or other elements.
A key structural motif is the benzene ring, a six-carbon cyclic molecule with alternating double bonds, exemplifying aromaticity. Benzene’s structure, first elucidated by August Kekulé in 1865, features delocalized pi electrons in a planar hexagon, conferring exceptional stability and resistance to reactions. This aromaticity arises from resonance: the electrons are shared equally, making the ring inert yet capable of substitutions. In biochemistry, benzene rings appear in aromatic amino acids—phenylalanine (Phe), tyrosine (Tyr), and tryptophan (Trp)—which contribute to protein folding via pi-stacking interactions and hydrophobic cores. They also feature in nucleotides: purine bases (adenine, guanine) contain fused rings including imidazole (five-membered) and pyrimidine (six-membered) components, akin to benzene’s stability. Benzene derivatives like phenols (from Tyr) play roles in signaling (e.g., neurotransmitters like dopamine) and antioxidants (e.g., vitamin E). In metabolism, rings enable energy storage, as in ATP’s adenine or chlorophyll’s porphyrin (a tetrapyrrole ring system with aromatic properties).
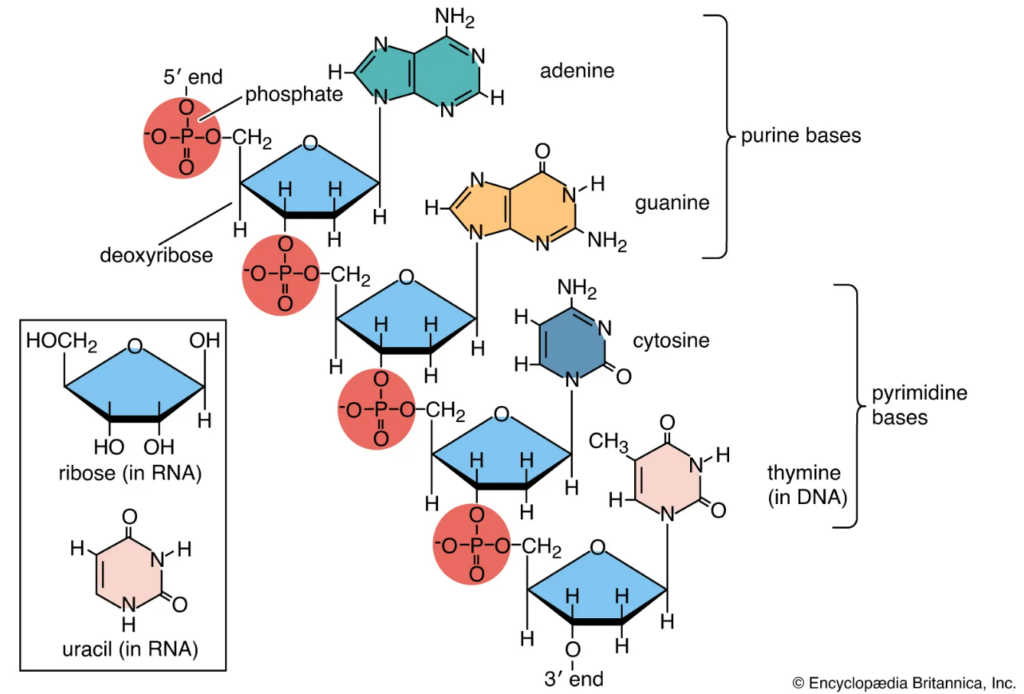
Nucleic acids—DNA (deoxyribonucleic acid) and RNA (ribonucleic acid)—store and transmit genetic information, sharing core similarities. Both are polymers of nucleotides, each comprising a nitrogenous base, a five-carbon sugar (pentose), and a phosphate group. The bases are purines (adenine-A, guanine-G) and pyrimidines (thymine-T in DNA, uracil-U in RNA, cytosine-C in both). These bases pair specifically: A with T/U (two hydrogen bonds), G with C (three bonds), enabling the double helix in DNA or single-stranded structures in RNA. The backbone alternates sugar-phosphate links via phosphodiester bonds, with bases projecting inward. This ATCG/U alphabet encodes genes via the genetic code, where triplets (codons) specify amino acids during protein synthesis.
A critical difference lies in the sugar: DNA uses deoxyribose, while RNA uses ribose. Ribose has a hydroxyl (-OH) group at the 2′ carbon of the pentose ring, making it an “oxy” sugar. Deoxyribose lacks this oxygen-containing group (hence “deoxy”), featuring just a hydrogen at the 2′ position. This seemingly minor change—DNA missing the 2′-OH—profoundly impacts stability. RNA’s 2′-OH enables hydrolysis (self-cleavage) under basic conditions, making RNA labile and suited for transient roles like mRNA or catalysts (ribozymes). DNA’s deoxyribose resists degradation, ideal for long-term genetic storage. This difference also affects structure: DNA forms stable B-form double helices, while RNA often folds into complex 3D shapes. Evolutionarily, RNA likely preceded DNA in an “RNA world,” with the 2′-OH aiding primordial catalysis before DNA’s stability enabled complex genomes.

Leave a Reply